2/06
Types of humoral immune responses and functional diversification of antibodies
B cell activation in antibody responses
division of labor among 3 mature B cell types
-
T cell dependent antibody responses
-
Follicular (Fo) B cells
- IgM + IgD
- most abundant
-
Follicular (Fo) B cells
-
T cell independent ab responses
-
marginal zone (MZ) B cells
- IgM
- about 1/10 of splenic B cells
-
B1 B cells
- IgM
- reside in fluidic parts of the body
-
marginal zone (MZ) B cells
T cell independent B cell activation
- there are 2 types:
- Type 1 - BCR activate B cells without T cell help, but using TLR / BAFFR ligand help
- Type 2 - BCR can activate B cells without T cell help, and uses no other help
review this
BCR can activate B cells with non-T help (TLR and BAFFR ligands)
-
TI type 1
- IgM aggregation + additional act sig
- simultaneous activation of BCR and other receptors on B cells (like TLRs) induce the B cells to proliferate and differentiate
- BAFFR - B cell activating receptor
- IgM + IgG
BCR can activate B cells without T cell help or other help
-
TI type 2
- IgM aggregation (extensive receptor aggregation)
- strong cross linking of BCRs by repetitive polysaccharide or protein epitopes
- IgM
role of innate immune signals in TI-type 1 B cell activation
CR2 complement receptor and CD19 are part of BCR as a whole
BCR do not function in isolation - have coreceptors
another component like complement can help even in T cell independent antibody response
important to keep in mind that T and B cells NEVER work in isolation
2 situations where have coreceptors -> help with B cell activation
- either with TLR or BAFF receptor
so:
- B cells not independent from T cells in function (know from before)
- and B cells can have help even when T cell independent - but still not as powerful as when T cell work with B cell
B cell Ab response requires signals in addition to Ag
need 2 signals in order to activate and undergo B cell proliferation + differentiation
- safe guard
- more tan 1 signal needed for activation, bc if not then easy for autoimmunity to occur
- so safe guard bc it keeps accidental self run from occurring bc need more than 1 signal to activate B cell
the germline center reaction
where T cell help is important bc changes in antibodies are much more potently helped by T cells
activation of B cells and migration of B/helper T cell complex into germinal center
-> B cell proliferation
-> somatic mutation and affinity maturation; isotope switching
-> exit of high affinity antibody secreting cells and memory B cells
sequence of events
- and 2.
- (steps 1 and 2): T and B cells recognize epitopes of the same antigen in diff locations
- T-B migrate to each other and B cell present antigen to T cells in the extra follicular space
- initially Ab are produced in the extrafollicular space
- activated Th and activated B cells migrate and form GC (germinal center) and are termed Tfh T cells and GC B cells
B cell can be anywhere in the body
B and T cell must have same specifications to work together to make potent immune response
must find each other for tangle to happen
this interaction (activation of B cell) not happening to germinal center (interaction occurs outside of germinal center)
make antibodies
bc new relationship, B cells make IgM isotype
antibodies produced help bridge gap btw initial exposure and have not refined binding site of antbodies
some T follicular and B cells move to germinal center (like discussed last class) which is important for maturation of B cells and memory cells
germinal center does not exist without infection
1. B and T cell activated independently
2. They find each other, B cell can start making antibodies early to cover the gap in time
differences in the antibody responses to TD (T cell dependent) vs TI (T cell independent) antigens
TD
- plasma cells made by T cell help can be long lived
- antigen = protein
- isotope switching
- IgM -> IgE, IgG, IgA
- yes affinity maturation
- secondary response (memory B cells)
TI
- plasma cells made independent of T cell help are short lived
- antigen = polymeric antigens, especially polysaccharides, glycolipids, nucleic acids
-
isotopes:
- TI2 = IgM
- TI1 = IgG
- little to no affinity maturation
- secondary response/memory B cells only with some polysaccharide antigens
polysaccharides conjugated to a protein carrier (TT) can recruit T cells and generate memory B cells
see pic
- bacterial capsular polysaccharide conjugated to protein recognized by B cell
- -> process the protein
- presents peptide to helper T cell
- B cell activation and differentiation
- -> long lived plasma cells and memory B cells
shows that can make T cell independent immune response into a T cell dependent immune response, bc B cell can attach to the pathogen (made of biomolecules)
complex of molecules has sugars, etc. so there is a mix of things, which B cell can bind bc they see shapes
what the B cell interact w/ gets internalized, and the protein in the pathogen gets processed by B cell, which is then presented to T cell -> now will produce long lived plasma cells (very used/handy in vaccine design, to provide longer time of resistance / better immunity)
they attach protein piece to a structure, can trick the T cell to help
What signals are triggered by TI type-2 versus TI type-1 T cell-independent antigens?
What is the stepwise process of T cell-dependent B cell activation and germinal center reaction?
pause for activity
Tfh-CD40L induces AID in B cells via CD40
see pic
from google:
Activation-induced cytidine deaminase (AID) is an enzyme that is predominantly expressed in germinal center B cells and plays a pivotal role in immunoglobulin class switch recombination and somatic hypermutation for antibody (Ab) maturation.
AID induces isotype class switching in B cells
Process where constant region can be switched to other constant region
Can switch constant regions without changing binding site
Why change binding site of antibody? So constant regions get switched
- each clone w/ same specificity has potential to change constant region, so have the same specificity
in the geminal centers: Ig isotope class switching
see pic
IgM - standard
if have T cell help: can switch to IgG, IgE, IgA
IgM - complement activation
IgG - Fc receptor dependent phagocyte responses; complement activation; neonatal immunity (placental transfer)
IgE - immunity vs helminths; mast cell degranulation (immediate hypersensitivity)
IgA - mucosal immunity (transport of IgA through epithelia)
overview of isotypes
- IgM, then can be switched into IgE, IgG, IgA
- framework: in T cell dependent immune response, the T cell produce cytokines, which influence what type of isotope is produced in the immune response
- initial pathogen which immune
response is made. The isotope made against it should be made to be
more effective vs the pathogen
- ex: infection in gut -> the immune response will make IgA, so isotope matches the pathogen type to help protection, bc it is in gut, so mucosal area which IgA is best for
primary antibodies can change their class by class switch recombination (CSR)
IgM - primary antibody
IgG - class switching CH
he didn't say anything
VDJ recombination creates binding site and class switch recombination (CSR) switches constant region of the Ab molecule
changing the DNA/genome of B cells to accomplish switch that constant region is changing
initially making IgM (constant region mu)
- this is what is normally made
same specificity can be juxtaposed to another constant region
constant regions can be switched and keep same specificity
HOWEVER, the genome itself obviously changes a LOT with these switches
Ig class organization in a naive B cell
see pic
initial transcription of VDJ recombination
turned out to be DNA recombination
Ig isotype class organization in a naive B cell
extremely long distances between Ig constant regions
longtranscripts or DNA recombination?
- turned out to be DNA recombination
Ig class organization in a naive B cell
DNA sequences before each constant regions can be joined together by an enzyme (cuts to where line goes)
result - same specificity now to make diff antibody isotype
T cell help w/ cytokines can each target DNA areas in chromosomes to make it available for recombination
if recombine to alpha, make IgA antibody, etc.
remaining DNA sequences are chopped out of genome, so then the isotope change is irreversible (permanent change in B cell clone)
fusion of 2 switch regions changes Ab class
rearranged DNA in IgM producing cells
-> signals from helper T cells -> AID
signal can make regions switch
carried out by AID
AID modifies DNA at the switch region to enable CSR
IgM -> AID
-> recombination os Su with Sy -> deletion of intervening C (constant) genes
-> transcription; RNA splicing
-> translation
= IgG
region not used -> deleted as a circle = now diff constant region
the intervening C regions are lost during class switch recombination
showing middle regions of C region being cut out during class switch recombination
already said this
in isotype switching, lose big chunks of genome
intervening DNA sequences of Ig CH regions are lost during CSR (class switch recombination)
same info
genes in heavy chain locus of an IgM expressing B cell
transcript for IgM
-> removal of DNA segment by enzyme activity between switch regions
-> non homologous end joining of DNA at switch regions
-> genes in heavy chain locus of an IgG expressing B cell
transcript for IgG1
SMH and CSR are 2 distinct processes
SHM (somatic hybrid mutation)
- in variable regions of gene (learned a long time ago)
and CSR
- at constant region of antibody
are BOTH carried out by the same enzyme, AID
V(D)J recombination generates antibody ---------.
SHM
increases antibody--------.
CSR switches antibody -------.
pause for activity
*******EXAM QUESTION!!!!*******
answer: diversity, affinity, constant region (isotype, IgG, IgA, etc.)
T cell cytokines determine CSR to distinct isotopes
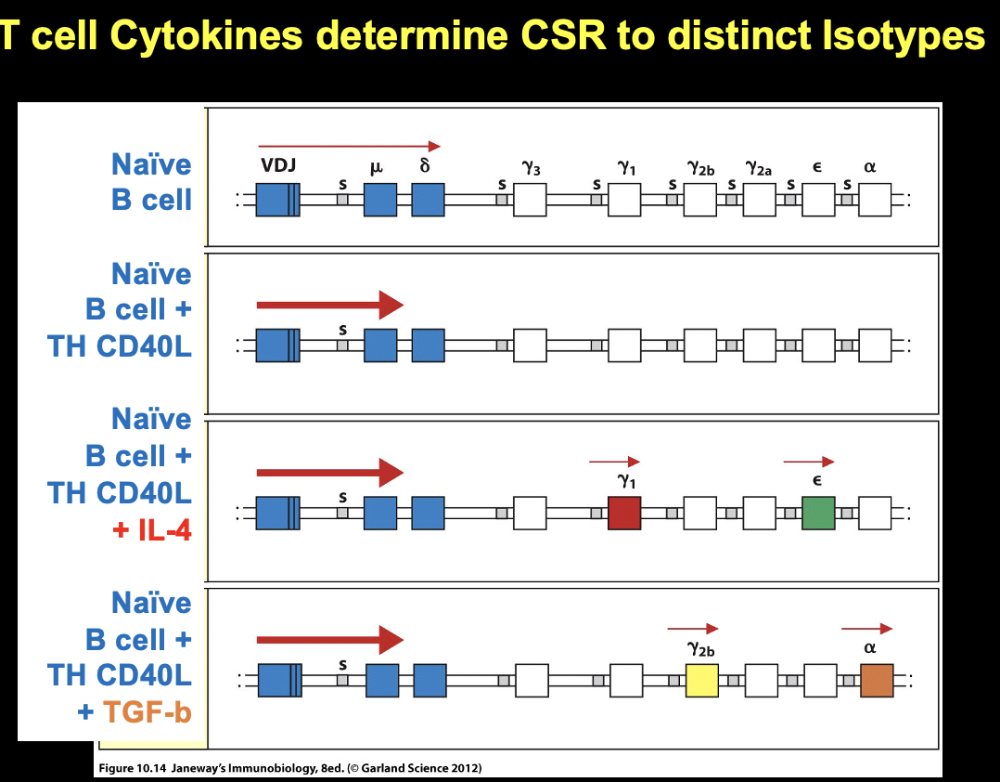
cytokines
- T helper cell can make IL4 - cytokine made by T follicular
helper cell can open constant regions
- cytokine specializes
in opening constant region Y1 (gG) and E (IgE)
- IgE important vs parasites and mucosal surfaces to protect skin
- IgG1 is good for most things
- cytokine specializes
in opening constant region Y1 (gG) and E (IgE)
- TGF-b opens Y2b and a locus constant regions
Ig heavy chain class (isotype) switching
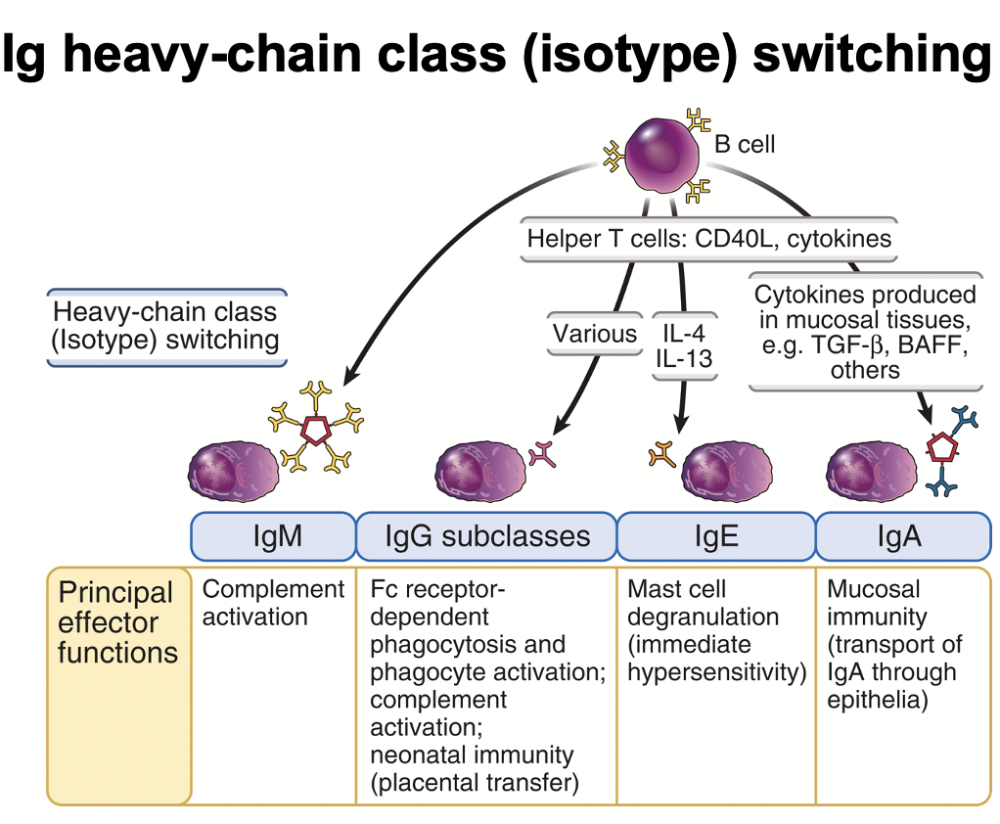
diff type cytokines made by helper cell allow switching to diff regions (have gone over this a lot already this lecture)
diff cytokines regulate the expression of diff antibody class
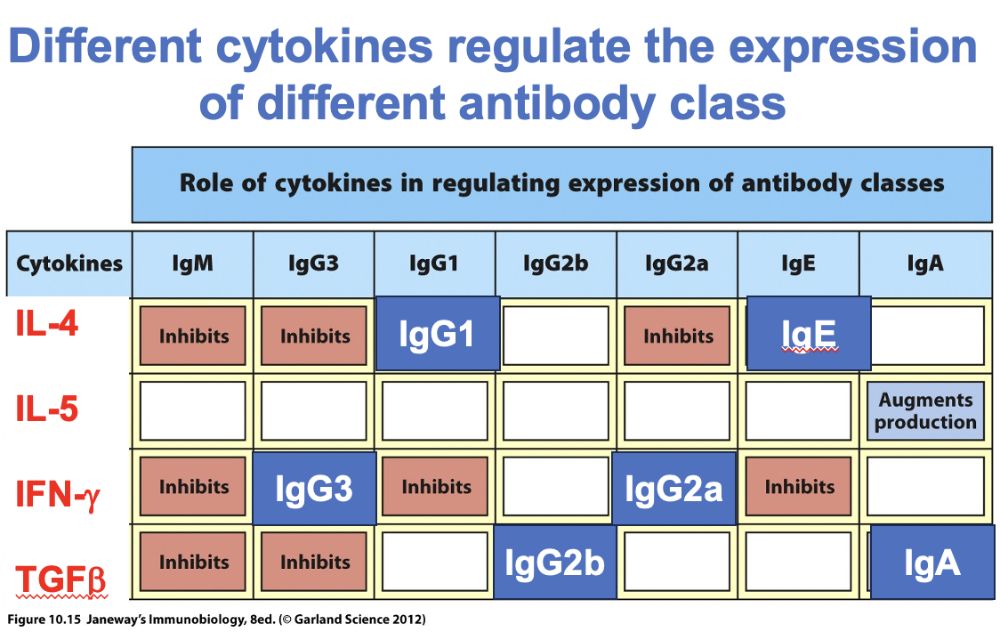
Summary to help remember
Which cytokines important for what
IL4 – IgE important for allergic rxns
Interferon gamma produced in immune response by T cells when pathogen is cross cellular
- result – antibodies produced help killing of target cells by T cells
Cytokines guide switching of antibody
major concepts in antibody generation + function
1. Exquisite specificity of Antibody for its cognate Antigen
- When antigen comes, choosing out of all b cell clones until fits lock and key
2. Adaptability of the Antibody to change its effector functions by changing Ig isotype by Class switch recombination (CSR)
3. Ability of antibodies to increase binding affinity (affinity maturation) by Somatic Hyper mutations (SHM)
- Can adapt to become better in binding by affinity maturation
4. CSR and SHM take place in specialized structures called Germinal Centers
- Geminal centers formed after B and T cell have been activated, meet, and form geminal center - before this is made, the B/T cell complex can make antibodies early which don’t generate memory (this is to fill gap btw innate response and when the better antibodies are made)
5. GCs are formed after T and B cells activated by the same antigen meet in extrafollicular spaces
6. GCs produce antibody-secreting plasma cells and memory B cells that produce recall antibody response upon reexposure with the same antigen (pathogen)
- Affinity btw antibody and antigen has tightest binding possible